This study was largely motivated by our collective frustration at the often inaccurate and sensational way that spiders are portrayed in the media. But just how bad is the quality of reporting on spiders, and why is it so pervasive?
To answer these questions and understand how misinformation about spiders flows at the global scale, we amassed a team of experts from all over the world to analyze 10 years of online news stories about human-spider encounters. We first searched Google News (using variations on the search term “spider AND bite”) to find more than 5000 news articles from 81 countries published in 40 languages. We then read each paper to collect data including the date and location of each reported event, whether the story described a human-spider encounter (but not a biting event) or a “bite” or “deadly bite” (spoiler: articles rarely included evidence of an actual spider bite), and checked each news item for errors (e.g., misidentified spiders in photos, incorrect information about spider biology or venom) and sensationalism (somewhat subjective**, but often based on the inclusion of words like “horror”, “terrifying”, and “deadly” ). We also recorded whether the article quoted an expert, and whether they were a medical professional, a spider expert (usually an arachnologist or entomologist), or “other” (e.g., a pest management professional). All of this data, detailed methods, and a summary of what we found broken down by continent is freely available online here.
An example of sensationalist reporting on a human-spider encounter in Australia.
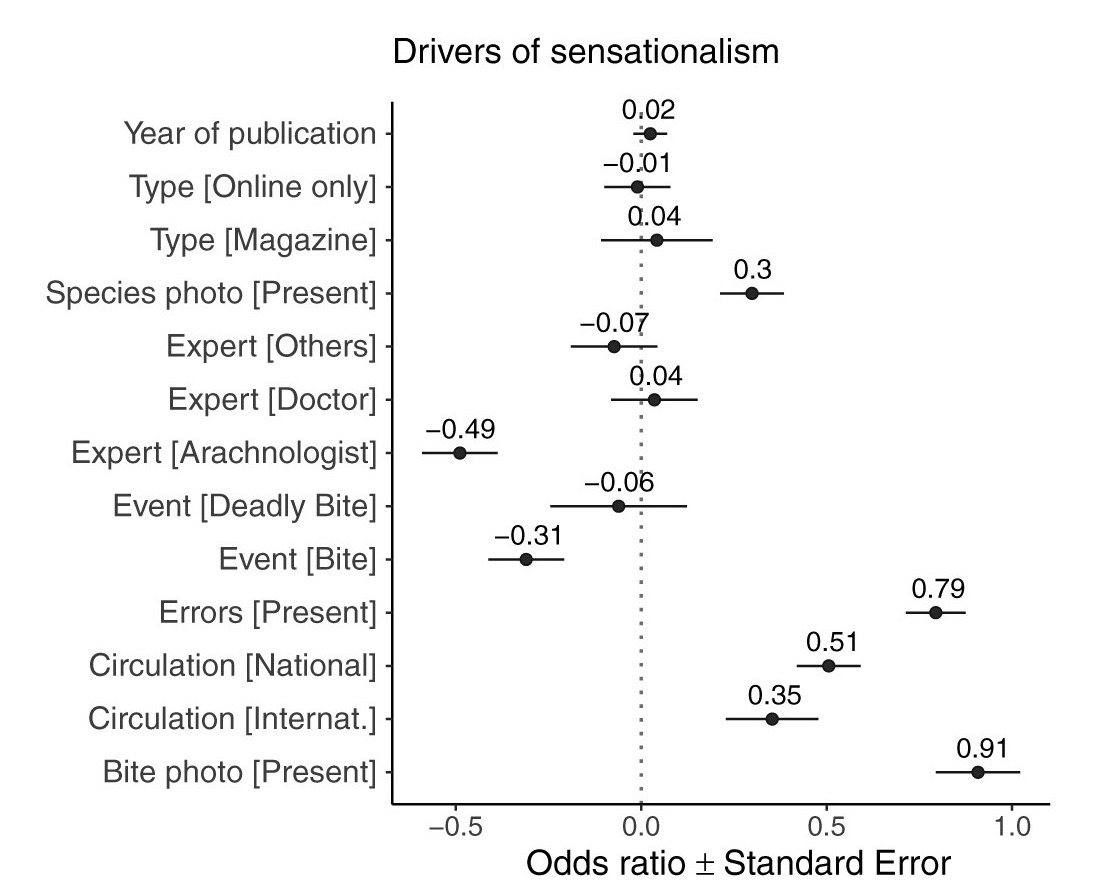
Overall, the quality of the reporting was poor: 47% of all articles contained one or more errors and 43% were sensationalist. Stories with photos of spiders or alleged bites were more likely to be sensationalized, as were stories that contained errors. Whereas quotes from medical or other experts were unrelated to sensationalism, stories that contained quotes from spider experts were much LESS likely to be sensationalized.
Drivers of sensationalism in media articles about spiders. Odds ratios to the left (right) of the dotted line indicate a decrease (increase) in the likelihood of sensationalism. Figure from Mammola et al. 2022.
One of our plans for the future is to create a global database of arachnological experts to make it easier for journalists to identify and contact arachnologists in their region who are willing to be interviewed and provide factual information about spiders. We are also already working on a paper outlining guidelines for journalists covering spider news stories, which we hope will help to prevent some of the most common errors we saw and improve the overall quality of reporting.
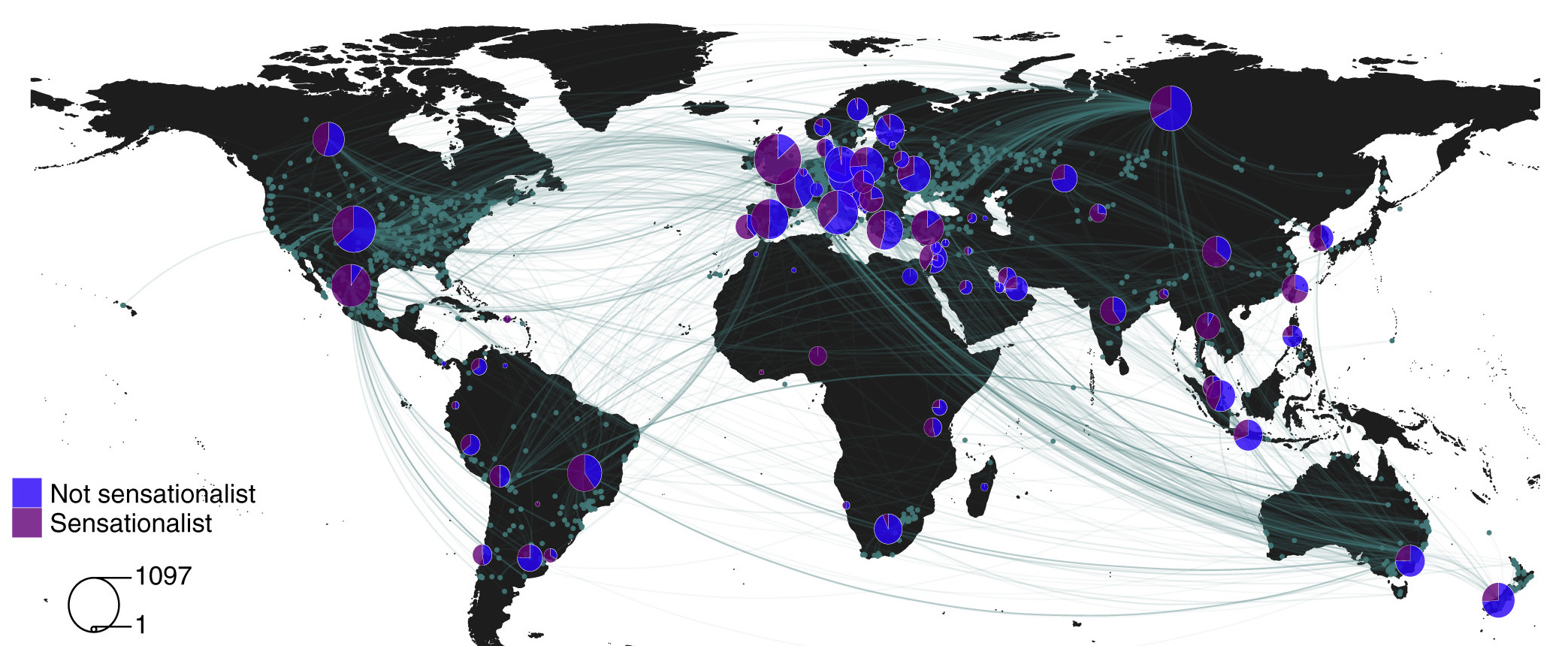
We next conducted an analysis to describe the flow of spider news stories around the world and to get at what may be driving the spread of (mis)information about spiders online. Unsurprisingly, countries with shared languages and with higher proportions of internet users were more likely to be connected in the global network. The number of medically important spider species present (i.e., those capable of harming and potentially killing humans) also increased the connectedness of individual countries within the network. Most notably, we identified sensationalism as a key factor underlying the spread of (mis)information.
Global distribution of news articles about on human-spider encounters showing links between countries (pies; n = 79) with each spider-related event reported by the press (dots, n = 2,644). The size of each pie chart represents the number of news articles published in the country between 2010 and 2020. Figure from Mammola et al. 2022
This study provides insight into what drives the global flow of information about spiders in particular, but can also teach us some more general lessons. Our results make us optimistic because they suggest a way to improve reporting on spiders, and in turn, to shift the quality and spread of online information more broadly. News stories are less sensationalized when they consult appropriate experts, and reducing sensationalism can help decrease spread of misinformation. We found that even local-scale events published by regional news outlets can quickly become broadcast internationally, which means improving news quality at the local scale can have positive effects that travel through the global network.
*Full author list: Stefano Mammola, Jagoba Malumbres-Olarte, Ingi Agnarsson, Valeria Arabesky, Diego Alejandro Barrales-Alcalá, Aimee Lynn Barrion-Dupo, Marco Antonio Benamú, Tharina Bird, Maria Bogolomova, Pedro Cardoso, Maria Chatzaki, Ren-Chung Cheng, Tien-Ai Chu, Naufal Urfi Dhiya’ulhaq, André-Philippe Drapeau Picard, Hisham K. El-Hennawy, Mert Elverici, Caroline S. Fukushima, Zeana Ganem, Efrat Gavish-Regev, Naledi Gonnye, Axel Hacala, Charles Haddad, Thomas Hesselberg, Tammy Ai Tian Ho, Thanakorn Into, Marco Isaia, Dharmaraj Jayaraman, Nanguei Karuaera, Rajashree Khalap, Kiran Khalap, Dongyoung Kim, Tuuli Korhonen, Simona Kralj-Fišer, Heidi Land, Shou-Wang Li, Sarah Loboda, Elizabeth Lowe, Yael Lubin, Marija Miličić, Alejandro Martínez, Zingisile Mbo, Grace Mwende Kioko, Veronica Nanni, Daniel Nwankwo, Yusoff Norma-Rashid, Christina Painting, Aleck Pang, Paolo Pantini, Martina Pavlek, Richard Pearce, Booppa Petcharad, Julien Pétillon, Onjaherizo Christian Raberahona, Joni A. Saarinen, Laura Segura-Hernández, Lenka Sentenská, Gabriele Uhl, Leilani Walker, Charles M. Warui, Konrad Wiśniewski, Alireza Zamani, Angela Chuang, Catherine Scott.
**For the news in English, Spanish, French, and Italian we checked to see how closely scores aligned for different collaborators assessing the same article, and we were pretty consistent.
This post is about a new paper just published online in the journal Animal Behaviour, titled “Web reduction by courting male black widows renders pheromone-emitting females’ webs less attractive to rival males” by Catherine Scott, Devin Kirk, Sean McCann, and Gerhard Gries! You can read the full text here (free access until 28 August, 2015). All photos and video are copyright Sean McCann.
This short video shows a male western black widow engaged in web reduction behaviour – a common element of the complex courtship rituals males perform on females’ webs. You can see him cutting some silk lines, then pulling silk out of his spinnerets with his last pair of legs, wrapping it around a bundled up section of the female’s web.
Web reduction behaviour is somewhat puzzling. This male black widow is attempting to convince a potentially cannibalistic female several times his size to mate with him. Destroying large areas of her home – which she relies on for both prey capture and protection from predators – is not the most obvious approach. So why do males do it (I know, the title of the paper probably gives this one away), and why do females let them?
Before we address the mystery of web reduction, let’s take a step back and set the scene for this story. It’s a pretty juicy one – there’s attraction, courtship, rivalry, and manipulation! Or maybe not manipulation… we’ll see!
Black widows are sexually dimorphic: the familiar female is on the left, and the much smaller and more brightly coloured male is on the right. My study species is the western black widow, Latrodectus hesperus.
Island view beach, on the Saanich Peninsula of Vancouver Island, BC, is a beautiful place to visit. The site shown below is on the lands of the Tsawout First Nation, who have kindly allowed me to collect spiders and do field work here over the past few years.
This coastal sand dune ecosystem supports a great diversity of organisms, but the black widows are the dominant web-building spiders. Females build their tangle-webs under the driftwood logs at a density of 2-3 webs per square metre of available habitat. There are many logs on this beach; correspondingly, there is a huge population of widow spiders.
At our field site, black widows build their webs under driftwood logs – often several females can be found living under a single piece of wood!
Female black widows almost never leave their webs, so when it comes time to mate, they need males to come to them. The solution to this problem comes in the form of pheromones on their silk. These chemical messages are kind of like scent-based personal ads, that provide the male with information about the female’s mating status and whether or not she’s well fed (sexual cannibalism is rare in this species, but starving females will sometimes eat males!).
If the female’s silk pheromone was actually a personal ad, it might say something like this!
Adult male black widows have only one goal in life: find a female to mate with (and become the father of as many offspring as possible). Once they mature, males stop capturing prey and abandon their webs for a more a nomadic lifestyle. If they detect an attractive female’s pheromone in the air, they follow it to her web.
A nomadic western black widow male traverses the sand dunes in search of a mate. Note: “pheromone trail” added for dramatic effect.
Once he arrives on the female’s web, the male begins an elaborate courtship display, dancing on the web to transmit vibratory messages(including “male, not meal!”) to the female. It may take many hours of courtship before copulation finally occurs.
Soon after he arrives, the male may start to destroy the female’s web. He cuts out sections, bundles them up, & wraps them in his own silk. Males usually reduce the area of the web by about 50%, and gather the destroyed sections up into loose silk-wrapped packages. These can be rope-like (as in the photo at the top of this page) or a tighter ball that has been wrapped extensively by the male like the one below.
Now to address our earlier question: why do the males engage in this behaviour?
Well, black widows are not the only spiders who do web reduction. Other species in the same family (Theridiidae) and others (including Linyphiidae) have similar behaviour. Watson showed in 1986 that web reduction in the sierra dome spider makes the female less attractive to rival males. He concluded that by bundling up the female’s web, the male decreases the surface area from which the silk-borne sex pheromone is released. Some researchers have assumed that it works the same way in other species. Others have suggested that web reduction could function in communication between the male and the female, perhaps improving vibration transmission through the web, or transmitting a male silk pheromone to the female. It could have one or both of these functions in black widows, but until now, no one had ever investigated!
The sierra dome spider, Neriene litigiosa (family Linyphiidae).
We set out to determine whether web reduction decreases female attractiveness to male black widows in a natural setting. We were pretty sceptical that reducing the web surface area by only about 50% could limit pheromone emission. Usually you have to decrease the dose of a pheromone by an order of magnitude to see any difference in attractiveness. However, males aren’t just bundling up the female’s pheromone-laden silk – they are also adding their own. We thought maybe it could be the addition of the male’s silk (and associated pheromones) that keeps other males away. We designed an experiment to try to find out.
First, we put a bunch of female spiders in cages and allowed them to establish webs. Then we loaded up the cages and took them, and a batch of males, to our field site.
This is what (part of) a truckload of black widows looks like. Here, my coauthor Devin is loading the cages containing females and their webs into the back of the lab pickup.
Once we got to our field site, we removed the female spiders from their cages (because we wanted to look at the attractiveness of silk only, not the females themselves) and set up four treatments: intact webs, mechanically reduced webs (with half the silk cut out), male-reduced webs (with about half the silk, on average, bundled and wrapped by the males we brought) and empty cages as controls (to confirm that captured males were actually attracted to the silk in the cages, not just wandering randomly).
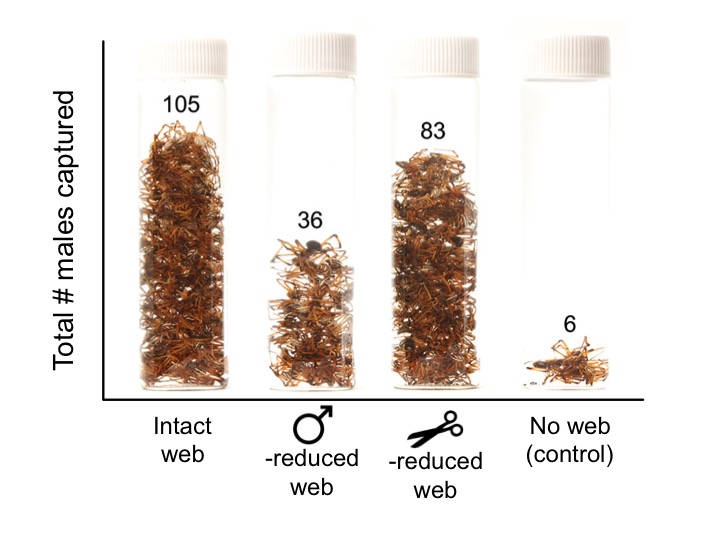
Four treatments: intact web, scissor-reduced web (50% of silk removed entirely), male-reduced web (courting males bundling up about 50% of silk on average), and no-web control.
We then turned the cages into traps that would capture any wild males attracted to the silk inside by surrounding them with sticky strips. Then we set the traps out on the beach in groups of four – one for each of the treatments.
The experimental setup. Each of the four traps contains a different treatment, and the white sticky strips surrounding the cages trap males that are attracted to the silk inside the cage.
We set the traps out at sunset (black widows are nocturnal) and waited to see what would happen. Soon the data started strolling in. We checked the traps every three hours, collecting and preserving any captured males. The sheer number of males out on the prowl was incredible – some webs attracted more than 10 males overnight!
Close-up of a trap containing a pheromone-laden female’s web, with a male black widow captured outside on the sticky strip.
After 24 hours, just by looking at the vials full of male spiders we had captured, the results were clear (if you want to see statistics, you can read the paper!). Male-reduced webs caught only about one third as many males as intact webs, so web reduction does in fact decrease attractiveness! As we suspected, however, removing half of the silk entirely did not significantly reduce a web’s attractiveness – we captured almost as many males outside scissor-reduced webs as intact webs. (A few spiders got trapped outside the empty control cages – they may have blundered into the sticky strips on their way toward an an attractive web.)
Beautiful data. It’s not often that the raw data tell the whole story, but here they do!
Evidently, when a male black widow reduces a female’s web, whatever he is doing is much more effective at decreasing its attractiveness than removing half of the pheromone-laden silk entirely. And he’s not actually removing any of the female’s silk – he’s just bundling it up into a ball. There are a couple of potential explanations for how web reduction works. Perhaps the female’s pheromone is not evenly distributed on the web, and the male targets the pheromone-rich silk for web reduction. Then, by wrapping those sections up in his own silk, he creates a barrier that limits the emission of the female pheromone. Another possibility is that the male’s silk has its own pheromone on it, one which other males detect and avoid. Or it could be a combination of both these mechanisms – we’re still not sure. We did another experiment to test the second idea, but the results neither supported nor completely ruled it out (see the paper for more details!). We will try to get to the bottom of this in the future.
Time for a cute male widow interlude! Look at him peeking out from behind that blade of grass.
For now though, let’s come back to the questions we set out to answer. Why do male black widows do web reduction? It allows them to monopolize the female, by making her web less attractive to other males. Courtship and mating last several hours, so if a male reduces the female’s web as soon as he arrives, he can decrease the likelihood of rival males arriving and interrupting. This may be very important at our field site, where competition for access to females appears to be fierce – during our second experiment, we had one intact web attract over 40 males in a single night! So web reduction is good for the male, because it helps him to avoid competition.
What about the female? Losing her web may be costly – she has to spend time and energy rebuilding it. However, we think she might actually benefit from web reduction too, and that the benefits may outweigh the costs. Sure, her web is important for prey capture and protection, but it’s also really attractive. So attractive, in fact, that even if she doesn’t add more pheromone, it will continue emitting its “come-hither” message for several days. Given number of males we saw arriving at each web during our experiment (40 in one night! even 10 is pretty extreme!), remaining attractive once she has already found a mate might not be so great. Having a choice between multiple males might be a good thing, but the female really only needs to mate once to fertilize all of her eggs. By “muting” her chemical signal though web reduction, the male might be doing her a favour: allowing her to rebuild her web without attractive pheromones (female sex pheromone production shuts off immediately after mating in black widows, but they don’t ever take down their existing webs). Rather than having to waste time and energy chasing off superfluous suitors, this may allow her to get on with the business of producing egg sacs!
Female western black widow guarding her egg sac. These spiders are very protective mothers!
References and further reading (also linked in the text)
The first paper from my MSc has just been published online in the Journal of Chemical Ecology! This study was a collaboration with colleagues Sean McCann (bioassay designer, photography/videography master, and all-around awesome assistant), Regine Gries (analytical chemistry wizard), Grigori Khaskin (synthetic chemist extraordinaire), and my supernatural supervisor Gerhard Gries. If you don’t have access to the journal, you can read the accepted manuscript here.
Here’s the story of the paper. Note: all photos and the video are copyright Sean McCann.
A female western black widow (Latrodectus hesperus) on her web. The silk is impregnated with sex pheromones that attract males and trigger courtship behaviour.
When I started my MSc, one of the goals for my research was to “find the pheromone” of the western black widow. What does that mean exactly? Well, we already knew that female black widows (spiders in the genus Latrodectus) produce sex pheromones that are somehow incorporated into the silk of their webs. These are sort of like chemical personal ads – they can provide information about things like the species, sex, age, mating history, and body condition of the individual producing them. When a male black widow matures, his only goal in life is to find a female to mate with. He abandons his web and follows his nose (not literally – we don’t really understand much about how spiders smell but their “noses” are most likely on their legs and pedipalps!) to a nearby female’s web. Given a choice among multiple available females, male black widows will go for a well-fed virgin based on the smell of her silk alone. Once he arrives at her web, he contacts the silk and “tastes” (again with receptors on his legs/pedipalps) the pheromone, which triggers courtship behaviour. We wanted to find out the chemical structure of the female’s sex pheromone.
We had a pretty good idea of what to expect, because other researchers had already identified a pheromone of the Australian redback spider (Latrodectus hasselti). It looks like this:
N-3- Methylbutanoyl-O-(S)-2-methylbutanoyl-L-serine methyl ester. Contact pheromone of Australian redback spider (Latrodectus hasselti) females.
Male western black widows are actually attracted to the webs of redback females, implying that the structure of the pheromone is similar, if not identical, in these two species. (It’s not necessary for males to discriminate between Australian redback and North American western black widow females in nature, because they never encounter one another, so it wouldn’t be that strange if they shared the same pheromone). So we set out to analyze the silk of our western black widow females, and see if we could find a similar compound.
We persuaded females to provide us with clean silk by allowing them to build webs on glass frames for three days. We then collected the silk and extracted it for use in behavioural experiments and chemical analysis.
Some silk collection, extraction, and analytical chemistry* ensued (I’ll leave it to you to read the paper for details if you’re interested), and just as we had hoped, our western black widow females had a compound on their silk that was very similar to the redback pheromone above:
N-3-Methylbutanoyl-O-methylpropanoyl-L-serine methyl ester. Candidate pheromone of western black widow females (Latrodectus hesperus).
Not only is this chemical similar to the redback pheromone, it is also present in small amounts on redback females’ silk. So it seemed like an ideal candidate for the western black widow pheromone, and provided a potential explanation for the attraction between the two species. Now all we had to do was make a synthetic version* of the pheromone and test it on actual males.
Before we could determine whether the compound we had found was in fact the pheromone we were looking for, we needed to come up with a way of comparing its effects with the real thing. We knew that contact with a female’s silk triggers courtship behaviour, but black widow courtship is long and complex, and involves several different kinds of behaviour, some of which are very subtle. The male’s courtship dance sends vibrations through the web to the female, possibly providing information about his quality and identity (including that he is a potential mate, not a meal!). It also involves the production of copious amounts of silk by the male. This male silk carries its own pheromones, and is deposited all over the web and onto the female herself in the form of a “bridal veil” during courtship.
Male black widow (L. hesperus) engaging in silk-wrapping on a female’s web during courtship. Here the male is wrapping a section of web that he has destroyed during web reduction behaviour, which I will discuss in a future post.
We designed an experimental setup to assess male responses to silk pheromones. We constructed this high-tech device out of bamboo barbeque skewers, laboratory labeling tape, and a paper cup filled with floral foam. The skewers form a “T” and at each end of the horizontal arm we slid on little envelopes made of squares of filter paper folded in half and stapled. This simple and inexpensive device was one of the big successes of the project.
Our simple and inexpensive T-rod for testing male behavioural responses to contact silk pheromones.
The T-rod design makes it easy to compare an individual male’s response to a test stimulus on one side (for example, a female’s silk wrapped around the paper envelope) to a control (blank paper) on the other.
Wrapping silk around a filter paper for behavioural experiments.
A male spider is introduced at the base of the “T” and climbs up to the top. Once he gets to the intersection, he can decide whether to go left or right. Males almost always began the experiment by investigating both sides of the “T”, but they spent much longer in contact with the silk-wrapped paper than the blank paper. Not only that, but they spent much of their time wrapping the female silk-wrapped paper with silk of their own – obvious courtship behaviour.
A male black widow silk-wrapping on a filter paper with silk extract on it.
Knowing that males would respond to female silk in this way on the T-rod, we were now ready to confirm that the behaviour didn’t depend on the structure of the silk itself, and to see if males would respond to our synthetic candidate pheromone in the same way as they would respond to the real thing.
We prepared female silk extract using methanol as a solvent (this is the same idea as vanilla extract, but instead of extracting the flavour of vanilla beans into ethanol, we extracted the chemicals on the silk into methanol) and applied it to one of the filter papers on the T-rod, and methanol alone to the other.
We tool silk from a glass frame like the one above and submerged it in methanol to extract the pheromones into the liquid, which we then used in behavioural tests.
Males responded in exactly the same way to silk extract as they did to silk itself, spending most of their time on the filter paper impregnated with extract, and wrapping it extensively with silk. Here’s a video showing what that looks like (first at full speed, and then slowed down):
This told us that a pheromone that can be extracted from the silk triggers courtship behaviour, and the structure of the silk itself is not necessary. But when we tested male responses to our candidate pheromone (dissolved in methanol, using methanol alone as a control), things were not so clear-cut. Males spent more time on the pheromone-impregnated paper than methanol alone, but they didn’t prefer it as much as they had preferred the silk extract to methanol. A few males engaged in silk-wrapping when they made contact with our compound, but not the majority, like we had seen for the extract. This means that although our “pheromone” elicited some male activity, by itself it is not enough to consistently trigger courtship behaviour. It seems to be a pheromone component – meaning that the pheromone is a mixture of one or more compounds in addition to the one we identified, and more work will need to be done to figure out what they are.
It would have been nice to be able to say we found the pheromone. But our results suggest that the chemical communication system of black widows is more complicated than we originally thought, and even more fascinating.
In the study that identified the redback pheromone, the researchers measured male activity (the amount of time they spent moving around when in contact with a filter paper impregnated with pheromone), not courtship behaviour. It could be that this pheromone too is only one component of a more complex chemical cocktail. Like our pheromone component, it may be responsible for eliciting searching behaviour, but not quite enough on its own to consistently trigger courtship behaviour by males.
If multiple compounds are involved in these spider pheromones, they might each have different functions. We don’t yet know whether the same pheromone that attracts males is responsible for triggering courtship, or if different compounds provide different kinds of information, about things like a female’s mating status and feeding history. We have learned that the scent-based sexual communication system of black widows is likely more sophisticated than we originally thought, and that there is much more to discover!
Male and female western black widow on a female’s web.
*Neither the analytical chemistry nor the synthesis of the candidate pheromone were trivial tasks – rather they required the expertise and generous efforts of my very talented coauthors Regine and Grigori. I hope they will forgive me for glossing over the details here!
Here’s the full citation for our paper:
Scott C, McCann S, Gries R, Khaskin G & Gries G. 2015. N-3-Methylbutanoyl-O-methylpropanoyl-L-serine Methyl Ester – Pheromone Component of Western Black Widow Females. Journal of Chemical Ecology. DOI: 10.1007/s10886-015-0582-x
This study is part of the PhD work of my friend and collaborator Samantha Vibert. In fact, we did some of the data collection and analysis for this paper during my very first semester in our lab, when I was working as an undergraduate research assistant. That was when I first began to really look closely at spiders and their incredible behaviour. My experience working with Sam that summer sparked my passion for the complexity and beauty of all of the various aspects of the private lives of spiders, which so often go unnoticed by humans.
Here is a plain-language summary of the paper, written with Samantha Vibert, and with photos by Sean McCann:
Spiders are fascinating but largely overlooked creatures, with sophisticated signalling systems involving chemical, vibratory, tactile, and in some species visual communication. A spider’s web is essentially an extension of her exquisitely tuned sensory system, allowing her to quickly detect and respond to vibrations produced by entangled prey. Not only is the web a highly effective prey-capture device, but it is also the dance floor on which prospective mates must demonstrate their desirability. The first moments after a male spider steps onto a female’s web may present a great risk, since spiders are often cannibalistic. We were interested in how a dancing male spider avoids a potentially deadly case of mistaken identity. One way that he might deal with this challenge is by transmitting vibratory signals that are very different from the vibrations produced by ensnared prey.
Spider webs are highly effective prey-capture devices, so how does a courting male avoid the fate of flies like this one?
Our study species were the western black widow and the hobo spider, which are both found in British Columbia.
A western black widow (Latrodectus hesperus) hanging from her tangle-web under a log at Island View Beach on Vancouver Island.
Black widows are in the family Theridiidae, and build complex, three-dimensional tangle-webs, while hobo spiders (family Agelenidae) build dense sheet-webs. Female black widows are much larger than males, while hobo spider males and females are closer in size.
A hobo spider (Tegenaria agrestis) female on her sheet web at Iona Beach, in Richmond, BC.
The purpose of our study was to describe some of the vibratory courtship signals of males in these two species, and to determine which aspects of these vibrations might allow females to discriminate between prospective mates and their next meals.
First, we recorded the vibrations transmitted through the web by courting males in both species using a laser Doppler vibrometer. At the same time, we video-recorded the male’s courtship behaviour. This allowed us to describe and analyze the different kinds of vibrations that were transmitted through the web during specific behavioural elements of each male’s courtship display. We then recorded the vibrations produced by the struggles of two types of common prey insects (house flies and crickets), on both black widow and hobo spider webs.
We found that male and prey vibrations differed more in the black widow than in the hobo spider. Hobo spider male vibrations contrasted with prey vibrations only in terms of their duration – the courting male moves around almost continuously on the female’s sheet web, while prey struggles are generally brief and intermittent. Black widow male courtship vibrations were also longer than prey vibrations on tangle webs (for the same reason), but they were also distinctive based on their generally lower amplitude and higher dominant frequencies.
To our surprise, we also found that most courtship behaviours in both species did not generate the kind of very stereotyped, complex and distinctive “songs” that have been reported in several other spider species. These species tend to court on substrates like leaf litter and plants, which most likely transmit vibrations quite differently than webs. Some male orb-weavers also produce highly rhythmic patterns during their vibratory courtship displays. So our finding leads us to wonder to what extent web architecture and complexity might constrain the transmission of the male courtship signals, and therefore the design of these signals.
One very interesting exception to the rule turned out to be the vibrations generated by the male black widow’s abdomen tremulations (an up-and down waggle of the abdomen, performed as the male hangs upside down from the female’s web). These vibrations were always very distinct from anything produced by prey: they were long-lasting and of very low amplitude, like a constant humming.
Here’s a short video of a male western black widow vibrating his abdomen on a female’s web (Supplemental File 1 from Vibert et. al 2014):
To learn more about these particularly stereotyped, ‘whisper-like’ male signals, we built our own custom web vibrator by modifying a loudspeaker. We were then able to play recorded vibrations of a male’s abdomen tremulation or a fly’s struggles back to females and observe their responses. Black widow females were much less likely to respond aggressively to vibrations played back at the “whisper-like” low amplitude of male abdomen tremulation, but attacked when we turned up the volume to levels typical of prey vibrations. This was the case regardless of which type of vibration we played. So we speculate that the males vibrate their abdomens either to avoid triggering a female’s predatory response, or even to turn it off.
Is it possible that the females that didn’t attack low-amplitude vibrations simply couldn’t detect them? We don’t think so. First, spiders are specialists when it comes to detecting even faint vibrations, and second, some females actually responded with courtship behaviour: abdomen ‘twitches’ which are similar to the male’s abdominal movements, but more emphatic. These abdomen twitches undoubtedly transmit their own vibrations through the web, and it would be very exciting to further investigate the female’s side of the vibratory ‘conversation’ during courtship.
Abdomen vibration seems to be a relatively common type of courtship behaviour and has been described in several spider families (‘abdomen wagging’ in an orb-weaver, and what has recently been described as ‘twerking’ in jumping spiders are a couple of examples). If indeed the “whispers” caused by these vibrations are involved in lowering female aggression, this might explain why such behaviour is fairly common among spiders.
The orb-weaver Argiope keyserlingi’scourthip also involves abdomen vibration, but in this species another vibratory signal was recently implicated in reducing the risk of cannibalism. The ‘shuddering’ of a courting male delays the female’s predatory response. One of the common features of black widow abdomen tremulation and these ‘shudders’ is that they are the first courtship behaviour performed by males after they enter a female’s web.
A male western black widow courting a large, potentially dangerous female. Abdomen vibration is performed on and off throughout the male’s courtship display, starting just after the male steps onto the web, and featuring prominently during attempts to approach and mount the female.
Very little is known about the kinds of vibratory courtship signals that male web-building spiders transmit to females through their webs, except for in orb-web weaving species. We hope that this new information about vibratory communication in tangle-web and sheet-web building spiders will contribute to better overall understanding of the function and evolution of web-borne vibratory courtship signals.
References:
Vibert, S., Scott, C., and Gries, G. (2014). A meal or a male: the ‘whispers’ of black widow males do not trigger a predatory response in females. Frontiers in Zoology, 11(4).doi:10.1186/1742-9994-11-4
Wignall, A. E., & Herberstein, M. E. (2013). The Influence of Vibratory Courtship on Female Mating Behaviour in Orb-Web Spiders (Argiope keyserlingi, Karsch 1878). PloS one, 8(1), e53057. doi:10.1371/journal.pone.0053057
Wignall, A. E., & Herberstein, M. E. (2013). Male courtship vibrations delay predatory behaviour in female spiders. Scientific reports, 3. doi:10.1038/srep03557
Mating behaviour and silk use in (some) mygalomorphs
So in my ‘intro to spider systematics’ post, I wrote that most of the Mygalomorphae are tarantulas. This is not actually true when you consider total numbers of species. Of the 2775 mygalomorph species (in 16 families), 950 species are in the family Theraphosidae (tarantulas), so that’s actually only about 34%. I probably should have said that the tarantulas are the most common mygalomorphs (and the ones people are generally most familiar with). Theraphosidae definitely wins the prize for the largest mygalomorph family, though. The next most speciose family is Nemesiidae, with 364 species.
I also implied that the Mygalomorphae aren’t all that sophisticated when it comes to silk use. Whereas the Araneomorphae have the ability to spin several different kinds silk specialized for particular functions, the Mygalomorphae produce only one general purpose silk. This is not to say, however, that they have not come up with some marvellous silk-based innovations!
In the centre of the vegetation in the foreground, you may spy the mouth of a well camouflaged silk-lined ‘turret’ built by a mygalomorph in the family Antrodiaetidae. (Photo by M. Hedin, licensed under CC BY 2.0)
Mygalomorphs have poor vision, so they must rely on chemical (smell or taste), vibratory, and tactile senses. It was traditionally thought that these relatively ‘primitive’ spiders had simple sexual communication systems. However, in their recent review in the Journal of Arachnology, Ferretti and coauthors argue that the sexual behaviour of mygalomorphs actually involves some quite elaborate courtship displays and complex mechanisms of communication. This post will highlight the 6 mygalomorph families described in the paper, most of which use silk in various ways for both prey capture and sexual communication.
Now, as a general rule, spiders are predatory, and mygalomorphs are no exception. They’re pretty keen to snap up anything that blunders into their field of vibratory perception, even if it’s another spider. The main functions of courtship behaviour in mygalomoprhs are mate recognition, orientation and synchronization of sexual behaviour, and suppression of non-sexual responses. Females must advertise their location and receptivity, and males must somehow signal “male, not meal!” to their potential partner. This can be tricky when the stage on which the male must show off his desirability as a mate is the same one that the female uses to detect prey. We’ll see what kinds of things these spiders get up to in order to make it to the finish!
Antrodiaetidae: turret mygalomorphs
Antrodiaetidslive underground in silk-lined burrows. The burrow entrance is extended with a turret made from silk, soil, and plant material that blends in beautifully with the surrounding substrate. The spider waits for prey just out of sight inside the tube, where she can detect and quickly respond to vibrations produced by insects brushing against the litter encasing the turret. Males detect the female’s location using pheromones – chemicals acting as olfactory personal ads – on the silk that lines her home.
Antrodiaetus riversi (Antrodiaetidae) in feeding position in turret (Photo by M. Hedin, licensed under CC BY 2.0)
Dipluridae: funnel-web mygalomorphs
The diplurid funnel-web is a horizontal silk sheet leading to a tubular retreat in a crevice or perhaps under a stone or log. From this silken hideout, the spider detects vibrations produced by prey passing over her sheet web. The female’s funnel-web also forms the dance-floor for the male’s vibratory courtship display. The male’s courtship signals are transmitted to the female through the silk sheet. Typically, she does not respond, which is great news for the male! Presumably his pedipalp drumming and many legged tap-dancing is quite distinct from prey vibrations, inhibiting the predatory tendencies of the female.
Ischnothele caudata (Dipluridae) female in her funnel web. Note the elongated spinnerets characteristic of this family. (Photo by M. Hedin, licensed under CC BY 2.0)
Mecicobothridae: sheet-web mygalomorphs
Mecicobothriidae is a family containing nine species of small spiders that build sheet-webs on the soil. In Mecicobothrium thorelli, chemical and/or tactile signals on the female’s silk trigger the male’s courtship display. Like in the diplurids, the male’s dance moves transmit vibrations through the silk to the female, who sits passively in judgment. The receptive female apparently enters a cataleptic state, allowing the male to haul her around the web and manoeuvre her into just the right position. It’s not quite as easy as all that for the male, though. In order for a successful copulation to occur, the chelicerae of the spiders must interlock in a very specific way. If the couple is disturbed while in copula, or there are any difficulties disengaging from this toothy embrace, it’s game over for the male, and dinner time for the female. Once the deed is done, a successful male will stick around on the female’s web, attacking any other males attempting to try their luck with the female. This ‘mate guarding’ behaviour is very unusual among mygalomorphs.
FemaleMecicobothrium thorelli(Mechicobothriidae) on her sheet-web. (Photo credit: Gabriel Pompozzi)
Theraphosidae: tarantulas
Theraphosids often live in silk-lined underground burrows, or silken retreats under rocks and vegetation. Sometimes they even build their tube-like homes in trees. Pheromones on the silk allow males to find and recognize females, and also trigger courtship behaviour. Males transmit species-specific vibratory courtship signals through the ground, to the female listening in her burrow. These seismic love songs can be detected by females at a distance of more than one metre. In Avicularia avicularia, the female actively responds, tapping her first pair of legs and pedipalps on the substrate. These good vibrations tell the male she is receptive to his advances, and may also help the male orient toward her.
Immature Avicularia avicularia (pinktoe tarantula) (Photo credit: Sean McCann)
Nemesiidae: tube-trapdoor mygalomorphs
Nemesiids generally live in silk-lined burrows, sometimes finished with hinged, camouflaged trap-doors. Male courtship includes scratching and tapping with the legs on the ground, often at a distance from the female’s burrow entrance, suggesting that this is another form of long-distance seismic communication. While most female mygalomorphs remain relatively passive throughout courthship and copulation, Acanthogonatus centralis females jerk violently, twitching all their legs and pedipalps. This enthusiastic behaviour may stimulate the male to begin copulation.
Calisoga longitarsis (Nemesiidae) at burrow entrance (Photo by M. Hedin, licensed under CC BY 2.0)
Microstigmatidae
The family Microstigmatidae contains 16 species of tiny spiders (males are only 1-3 mm long!). This is one family that apparently makes minimal use of silk. Xenonemesia platensis males only begin courtship after making direct contact with the female’s body. The male’s courtship behaviour includes quivering with the first two pairs of legs. The female, if she is receptive, responds by moving into a mating posture with her genital area exposed, allowing the male to clasp her pedipalps and chelicerae with his first legs.
This clasping of the female’s chelicerae by the male is common in mygalomorphs; in many species males have specialized structures on their legs that facilitate the embrace. Some researchers think it may be a way of restraining a potentially lethal female, keeping her fangs at leg’s reach. However, the fact that female catalepsy during copulation is also widespread sheds some doubt on this interpretation. Alternatively, it may be a more ‘symbolic’ form of bondage akin to the bridal veil in some araneomorphs. The tactile stimulation associated with the male’s clasping may in fact cause the female to become quiescent.
Throughout copulation, the Xenonemesia platensis male continues his tactile courtship, tapping and scraping his second pair of legs against the female. It’s possible that this copulatory courtship persuades the female to use his sperm over that of competitors, but more work needs to be done to test this hypothesis.
Mating Xenonemesia platensis (Microstigmatidae). The male has the female’s pedipalps and chelicerae clasped in his first pair of legs, while he uses his second pair of legs to beat and scrape the female’s coxae (leg segments closest to the body) (Photo credit: Gabriel Pompozzi)
The mygalomorphs may all look fairly similar, but they have surprisingly diverse habits, and use their multi-purpose silk in clever ways. Their sexual communication is most certainly not simple, incorporating multiple signaling modalities. Just as with most araneomorphs, good vibrations and sexy scents are the key!
Growing evidence suggests that contact sex pheromones associated with female silk are common in mygalomorphs (as they are in araneomorphs). Sex pheromones have also been implicated in the sexual communication systems of some spiders in the Mesothelae, suggesting that chemical communication was acquired early in the evolutionary history of spiders.
This story of mygalomorph mating also highlights the fact that female spiders in general are not simply passively waiting for marauding males to stumble across their doorsteps. On the contrary, they actively advertise for a mate, sometimes participating in a vibratory signaling ‘conversation’ with courting males, and likely judge a suitor’s quality both before and during copulation.
The private lives of spiders never cease to fascinate!
Special thanks to Gabriel Pompozzi for allowing me to use his photographs of Xenonemesia platensis and Mecicobothrium thorelli. Check out more of his fantastic images of mygalomorphs here.
I am also very grateful to Marshal Hedin for generously making his photographs available under creative commons licenses. He has some really wonderful sets of shots of several mygalomorph families on flickr.
Following from last week’s story about silk bridal veils, this post focuses on another rare use of silk in spider courtship behaviour: the giving of silk-wrapped ‘nuptial gifts’.
This phenomenon has been most well studied in two spider species in closely related families: Pisaura mirabilis (Pisauridae)
Pisaura mirabilis male carrying a silk-wrapped nuptial gift. (Photo by Ferran Turmo Gort, licensed under CC BY 2.0)
These spiders can teach us three valuable lessons about gift-giving in advance of the holiday season*.
1. Gift-giving can improve mating success.
In both families, nuptial gift giving behaviour is essentially the same. Before mating, the male obtains a prey item, (usually) wraps it up with silk, and offers it to the female during his courtship display. If she’s in the mood, she’ll grasp the package in her chelicerae, and while she’s busy consuming the prey inside, the male will copulate. It’s possible to mate without providing a gift, but in both Pisaura mirabilis and Paratrechalea ornata, males that give gifts have higher mating success: they have longer copulation durations and fertilize more of the female’s eggs.
2. Failure to wrap delicious gifts may result in their consumption, but no sex.
Females willingly accept unwrapped prey items, but may run off with them before mating can occur. This happens much less frequently with wrapped gifts, which are easier to hang on to. In Pisaura mirabilis, if the female tries to abscond with a gift that he still has in his grasp, the male goes limp (called thanatosis, or death-feigning) and allows himself to be dragged along with the gift. Once the female settles down to eat it, the male springs back into action and copulates.
3. Attractive silk gift-wrap will effectively disguise useless items, but only for a limited time.
Visual appeal may play some role in whether females accept nuptial gifts, but in Paratrechalea ornata, there are chemical cues specific to the male’s gift-wrapping silk that elicit female grasping behaviour. One of the advantages of this is that males can get away with giving females worthless items such as seeds, plant material, or prey that they’ve already fed on. Provided it’s wrapped up in attractively scented (or tasty) silk, the female will accept the gift and the male can copulate. As soon as the female realizes there’s no food inside the package, however, she’ll cut the mating short.
So there we have it. Pick the perfect prey item, wrap it up in silk, and hang on tight!
*Lessons may have limited application to non-spider interactions
I’m currently sifting through mountains of literature on spider biology searching for references to silk use in courtship and sexual communication.One of the particular topics I’m interested in is the rarely reported ‘bridal veil’. So far I’ve found records of bridal veils in 12 families, all in the Araneomorphae. I’ve included photos of most of the species in question (or a species in the same genus) to highlight the morphological diversity of the spiders that share this weird and wonderful behaviour!
Safe sex
Courting a female can be a risky business. Spiders are predators (with some exceptions), and sometimes highly aggressive females would rather treat a male as dinner than a date. Male spiders have diverse, complex repertoires of courtship behaviours, some of which most likely function to inhibit the predatory tendencies of females.
The paired pedipalps of male spiders are modified for transferring sperm. This means that, usually, the male needs to copulate twice in order to secure paternity of as many offspring as possible. The mechanics of copulation are often complex, and the male can’t afford any untimely interruptions. Possibly, the bridal veil has a role to play…
Getting tied down
Bristowe coined the term ‘bridal veil’ in his 1958 paper describing the mating behaviour of the crab spider Xysticus cristatus and Xysticus krakatuensis (Thomisidae).
Xysticus cristatus female (photo credit: Arlo Pelegrin)
Part of the male’s courtship behaviour includes anchoring the female’s legs and cephalothorax (front body segment) to the substrate with a ‘veil’ of silk threads. After mating, the female apparently has no trouble freeing herself from her silken bonds.
The centimetres high club
The nursery web spider Pisaurina mira (Pisauridae) puts a spin on the bridal veil idea.
In this species, courtship and mating take place as the spiders hang in midair, suspended by their draglines below a leaf. Before copulation, the male ties up the female’s first two pairs of legs in front of her cephalothorax, by spinning silk as he twirls the female around on her thread. Bruce and Carico (1988) suggested that the split-second that it took for the potentially cannibalistic female to struggle free from the veil gave the male just enough time to climb up out of harm’s way.
Oxyopes schenkeli (Oxiopidae)males have very similar bridal veiling behaviour, which results in the binding of the female’s first three pairs of legs with silk.
The context of courtship in these spiders is also up in the air, suspended by silk draglines from a leaf (60-90 cm above the ground). After observing at least one male getting cannibalised despite spinning a bridal veil, Preston-Mafham 1999proposed that the main function of the bridal veil is to stimulate the female to mate, possibly via pheromones (chemical signaling molecules) on the silk.
A touch of silk
The courtship of Dictyna volucripes (Dictynidae), takes place on the female’s web.
The male begins by depositing silk on the web, at a distance from the female, before approaching and applying a light silk wrapping to her body. Starr (1988) concluded that males of this species are not in any real danger from females – although females occasionally rushed towards males, males were able to easily avoid them.
Meta segmentata is a long-jawed orb-weaver(Tetragnathidae).
The veiling behaviour in this species was described as “partial wrapping of the female as though she were prey”. Lopez (1986)suggested that the silk of the bridal veil might inhibit female aggression through physical contact with sensory hairs on her body.
Throughout copulation, Schizocosa malitiosa (Lycosidae)males release dragline silk over the upper surface of the female’s front legs.
Penultimate (one molt away from maturity) Schizocosa male (photo by Marshall Hedin, licensed under CC BY 2.0)
A fairly sparse bridal veil is a common element of courtship in the genus Latrodectus (Theridiidae).Since I study western black widows (L. hesperus), I’ve included a video of one of ‘my guys’ doing his thing (video taken by Samantha Vibert)
Ross and Smith (1979), studying L. hesperus, and Aisenberg et al.(2008), studying S. malitiosa,suggested that the bridal veil silk is impregnated with a pheromone that induces female catalepsy. Placing the pheromone-laden silk directly on the female’s body might be the best way to ensure that she receives the chemical message and remains passive throughout copulation.
Homalonychus theolougus penultimate male (photo by Marshall Hedin, licensed under CC BY 2.0)
These are wandering spiders, and mating takes place on the ground. With the female’s legs all drawn up close to her cephalothorax, the male circles around her, binding her legs together tightly with a ring of silk. After the first copulation, he’ll add some more silk, reinforcing the ring, then mate a second time. As soon as the second copulation is completed, the male beats a hasty retreat. A second later the female breaks free from the silk ring, and spends some time grooming, trying to remove all the silk from her legs.
Thalassius spinosissimus (Pisauridae) females build a special mating web and hang from it in ‘mating posture’ with all the legs drawn in tightly as described above for the homalonychids. Males in this species also ring the female’s legs with silk (Sierwald 1988).
The male starts by spinning an ‘outer’ ring of silk around the female’s tibiae, then he adds a second, ‘inner’ ring around the patellae (see diagram with names of leg segments here). His handiwork complete, he tips the trussed-up female over onto her side and mates with her (Merrett 1988).
Cupiennius coccineus (Ctenidae)males, in staged encounters with heterospecific (Cupiennius salei) females, sometimes engaged in bridal veil spinning behaviour.
Here’s where it starts to get interesting. Normally, when mating with females from their own species, Cupiennius males don’t go in for the bridal veil thing. However, when researchers paired C. coccineus males with C. salei females (who are on average a bit bigger than the C. coccineus females), some males circled the female, depositing silk on her legs. Two of the three males that spun bridal veils were able to mate with the heterospecific females, while the third became lunch. As Schmitt (1992) noted in reference to this unfortunate male’s demise, “Obviously, the male silk did not seriously affect the female’s mobility.”
One explanation is that the ‘veil’ in this situation is a result of some confusion over whether to treat the too-large female as a potential mate, or prey (these guys can take down prey larger than themselves and normally use silk in this context). Another option is that this is a part of the courtship repertoire of Cupiennius males, but it’s reserved for especially large, potentially dangerous, females and was never seen before because usually similarly sized individuals were paired for laboratory mating observations (Schmitt 1992).
Courtship in both Argiope aemula (Araneidae),
Argiope aemula female (photo by falilin, licensed under CC BY 2.0)
and Nephila pilipes (Nephilidae),
Nephila pilipes female (photo by drriss, licensed under CC BY 2.0)
takes place on the female’s orb-web. The tiny male does a variation on the ring-type bridal veil, doing his silk spinning on the top of the female’s cephalothorax and abdomen (he’s so small he has room to walk around on there). He attaches silk at the bases of the female’s legs, building up a complex network of silk (Robinson and Robinson1980).
Lifting the veil
Recently, Zhang et al. (2011) published the first experimental study of the function of a bridal veil. The authors wanted to figure out if the bridal veil in Nephila pilipes has any role in reducing female aggressiveness, and if so, whether chemical and/or tactile cues were responsible.
In the lab, males never spun bridal veils prior to their first copulation. When female movement interrupted the first copulation, males that deposited silk inevitably copulated a second time, while most males that tried to mate again without spinning a bridal veil were cannibalized.
The researchers then compared the success of normal, silk-slinging males with males that had their spinnerets covered with super-glue*. It turned out that these males did just as well as normal males by going through the motions of bridal veiling behaviour even though they were prevented from spinning silk. Further experiments preventing females from detecting potential chemical and/or tactile cues associated with bridal veil spinning behaviour suggested that both touch and smell are likely involved.
Tying it up
Are bridal veils physical restraints or stimulating strands? Is silk a substrate for sexy scents or catalepsy-inducing compounds? It’s really not clear. Given that female spiders commonly produce silk-bound pheromones, I suspect that male silk pheromones are probably important. However, it’s becoming increasingly apparent that spider communication systems are highly sophisticated, and messages may be simultaneously transmitted between individuals via multiple modalities. Spiders use vibratory, chemical, tactile, and (sometimes) visual signals and senses in a variety of combinations, and untangling this mystery will take a lot more investigation!
*In case you’re concerned that super-gluing spiders is not a nice thing to do, I can assure you that cyanoacrylate is recommended for use on spiders in the book Invertebrate Medicine. I’ve looked into it because I’ve done some spider gluing myself.
References without direct links in the text:
Bristowe, W.S. 1958. The World of Spiders. Collins, London.
Lopez, A. 1986. Glandular aspects of sexual biology. In: Ecophysiology of Spiders (Nentwig, N., ed.). Springer Verlag, Berlin, pp. 121—131.
Merrett, P. 1988. Notes on the biology of the neotropical pisaurid, Ancylometes bogotensis (Keyserling) (Araneae: Pisauridae). Bulletin of the British Arachnological Society. 7: 197-201.
Preston-Mafham, K.G. 1999. Notes on bridal veil construction in Oxyopes schenkeli Lessert, 1927 (Araneae: Oxyopidae) in Uganda. Bulletin of the British Arachnological Society. 11(4): 150-152
Schmitt, A. 1992. Conjectures on the origins and functions of a bridal veil spun by the males of Cupiennius coccineus(Araneae, Ctenidae). Journal of Arachnology 20:67–68.
Acoustic communication is common in spiders, but the vibrations are usually transmitted through a substrate, like the ground, leaves, or webs. Spiders detect substrate-borne vibrations with highly sensitive receptors on their legs. In other words, spiders don’t have ‘ears’ or hear in the way that humans do, while we are generally deaf to the sorts of acoustic communication signals they are sensitive to. As a new tweeter, I was excited that I could answer this question, having recently read a book chapter by Gabriele Uhl and Damian Elias on spider communication. They mentioned a tarantula called Theraphosa blondi, or, commonly, the Goliath Birdeating Spider (don’t worry, more about this intriguing name later) producing a defensive hissing sound like a snake. Importantly, this audible (to humans) sound occurs during communication with vertebrate predators, rather than other spiders. Recently, I read the original paper by Marshall et al. (1995) describing the mechanism of sound production by Theraphosa blondi, which will be the topic of this post.
First off, a little background on the ‘Goliath Birdeaters’. There are two species: Theraphosa blondi (or leblondi, depending where you look) and Theraphosa apophysis. I had the pleasure of meeting large females of both these species while in French Guiana doing some field work on army ants with Sean last winter.
Female Theraphosa blondi in French Guiana. A quick spider anatomy lesson: spiders have two body segments – the front one with the legs is the cephalothorax (sometimes called the prosoma) and the rear one is the abdomen (or opisthosoma). In addition to 4 pairs of legs, the front-most pair of appendages are modified legs called pedipalps (sometimes just palps). Notice the hairs on the rear of the abdomen – these are the defensive urticating hairs mentioned later.
The Goliath Birdeaters live in burrows. Tarantulas (members of the family Theraphosidae) don’t build webs, but here you can see the silken ‘doormat’ that extends from the silk-lined burrow of this T. blondi female. We tried to lure her out by twisting a twig around on the silk mat at the burrow entrance.
The spider detected the vibrations from the movement of our stick through her legs, which are always in contact with the silk lines. Presumably we did a passable job of imitating a prey item, as she soon rushed out of the burrow to attack, biting the stick. Her fangs were impressive (about 2 cm long) but the venom of these spiders is not that potent – a bite is apparently not much worse than a wasp sting.
Female Theraphosa apophysis outside her burrow. This spider has evidently seen some rough times, having lost her last two legs on her left side.
The ‘goliath’ part of the name is appropriate: they are extremely large.
A Theraphosa blondi female next to a woman’s hand for scale.
As for ‘birdeater’, like most spiders, they mainly prey on insects and other arthropods. They have also been reported to take juvenile toads, skinks, snakes, mice and earthworms, but rarely birds*.
Spiders have predators themselves, which is where defense mechanisms come in handy. Tarantulas in the New World employ urticating hairs in defense against predators such as small mammals. These barbed bristles on the spider’s abdomen are brushed off with the last pair of legs in the direction of an attacker and can result in severe irritation of the skin and mucosa. Coatis (members of the raccoon family) prey on tarantulas and apparently use tactics that allow them to avoid being injured by their prey’s urticating hairs.
White-nosed coati (a relative of raccoons in the family Procyonidae). These guys like to eat tarantulas. (Photo courtesy Adam Blake)
Coatis have a severe reaction to the hairs of Theraphosa blondi, however, and seem to recognize and avoid them. The authors of the paper suggest that the hissing sound the spiders produce is an acoustic aposematic (warning) signal that works in combination with the urticating hairs to provide an effective defense against vertebrate predators. The point of their study was to figure out exactly how these giant spiders hiss.
When the researchers looked closely**, they found that the spiders have many plumose setae (tiny hooked hairs) on the femora of their pedipalps and first two pairs of legs. When threatened, Goliath Birdeaters rear up and rub these leg and pedipalp segments together.
Theraphosa apophysis. The arrow indicates the femur of the right first leg. The plumose setae referred to in the paper are on this leg segment as well as on the femora of the pedipalps and second pair of legs.
Experimentally removing the setae completely silenced the spiders during their threat displays. Sequentially removing more and more of the hairs caused the amplitude of the hissing sound to decrease incrementally, but the frequency remained the same. Using dead or anaesthetized spiders, it was possible to produce pretty much the same sound by rubbing the legs together the way the spider would. The authors concluded:
“The mechanism of sound production is apparently the entanglement of the terminal hooks of one seta with the filaments of the plumose setae on the opposing leg surface. This produces sound in a similar way (and with similar effect) that hook-and-loop fabric closures (marketed as Velcro) do during opening.”
So basically, although Goliath Birdeaters don’t really eat all that many birds, they have a nifty trick for avoiding being eaten themselves. They use a velcro-like technology to make a hissing sound that warns potential predators that they are dangerous and painful to attack. Pretty awesome.
*I haven’t been able to track down any references for this, but I have read in a few places that the original species description for Theraphosa leblondi included a record of the spider having taken a hummingbird.
**The figures in the original article Marshall et al. 1995 (paywalled) include close-up images of theses structures. For (non-paywalled) images of stridulatory setae in another Theraphosid see Pérez-Miles et al. 2005.






















































{kind=link}